Pterosaur flight
This is a work in progress; updates will occur occasionally.
Take-off and wing structure

(Image of Tupuxuara taking off by Julia Molnar; note the muscle fibers of the patagium are contracted, making the membrane thinner, something that'd probably help in terrestrial locomotion)
For a pterosaur, the first step would be leaving the ground. Unlike birds, which, as bipeds, have strong hindlimbs, which propell the animal as it leaves the ground, pterosaurs had stronger forelimbs, the arm/wing bones; this implies they used them instead to take the animal off the ground, for the hindlimbs would be too weak for this. Much like modern vampire bats, pterosaurs seemed to have used a catapult like motion of the front limbs (a frog leap using the hindlimbs before using the front limbs could give aditional power), which would certainly be more powerful than in bats thanks to the long metacarpals. This technique clearly is more efficient than the one used by large birds, since it wastes far less energy than running, and presumably allowed pterosaurs to reach larger sizes. Also this means that the hindlimbs do not need to be particularly robust and that no heavy musculature on them is needed, as opposed to the useless weight birds have to carry on the air. This method most likely used lots of anaerobic respiration, specially on pterodactyloids, which have long necks, creating a dead air space, as well as smaller bodies and therefore smaller lungs. Both ornithocheiroid and azhdarchoid pterosaurs (incidently the ones that got really big) developed air sacs in the patagia along the front limbs, perhaps as means to suply the muscles with oxygen imediately after take off.
Once in the air, a pterosaur would fly with what can be called the most complex wings of the animal kingdom. The pterosaurian wing basically consisted of a membrane stretched along the arm, long metacarpals and even longer fourth finger, connected to the body and backlimbs; a smaller membrane, the propatagium, extending from the shoulder to the wrist (which had a long, thin bone called pteroid) and probably along the metacarpals, and another membrane between the legs (the uropatagium, which we will debate further). The propatagium and the uropatagium were simple membranes, but the main patagium, the cheiro/brachiopatagium, was very complex; it had several, extremely thin layers inside, composed of muscle fibers and collagen fibers (actinofibrils); supplying these layers were blood vessels, which unusually had loops on them. These fiber layers had several purposes; the muscle fibers imply the animal could extend and contract the membrane, thus changing the shape of the wing, making it thinner or wider. The actinofibrils were used to make the membrane more stiff, as mere skin would be too weak for flapped flight (bats do have wing fibers, though they are not as specialised as pterosaurian ones); being more concentrated on the distal part of the wing, they made the wing tips more rounded, as opposed to old reconstructions of pointy winged pterosaurs, as pointy wings would result in more falling than flying. As mentioned above, some pterodactyloids had air sacks on the wings; inflating them would probably help making the wing wider, though the main purpose was to pneumatise the forelimbs and perhaps supply the wing muscles with oxygen.
[Below, a picture of the Kitakyushu pterosaur by Matteo Bachin; note the rounded wing tips)
For a pterosaur, the first step would be leaving the ground. Unlike birds, which, as bipeds, have strong hindlimbs, which propell the animal as it leaves the ground, pterosaurs had stronger forelimbs, the arm/wing bones; this implies they used them instead to take the animal off the ground, for the hindlimbs would be too weak for this. Much like modern vampire bats, pterosaurs seemed to have used a catapult like motion of the front limbs (a frog leap using the hindlimbs before using the front limbs could give aditional power), which would certainly be more powerful than in bats thanks to the long metacarpals. This technique clearly is more efficient than the one used by large birds, since it wastes far less energy than running, and presumably allowed pterosaurs to reach larger sizes. Also this means that the hindlimbs do not need to be particularly robust and that no heavy musculature on them is needed, as opposed to the useless weight birds have to carry on the air. This method most likely used lots of anaerobic respiration, specially on pterodactyloids, which have long necks, creating a dead air space, as well as smaller bodies and therefore smaller lungs. Both ornithocheiroid and azhdarchoid pterosaurs (incidently the ones that got really big) developed air sacs in the patagia along the front limbs, perhaps as means to suply the muscles with oxygen imediately after take off.
Once in the air, a pterosaur would fly with what can be called the most complex wings of the animal kingdom. The pterosaurian wing basically consisted of a membrane stretched along the arm, long metacarpals and even longer fourth finger, connected to the body and backlimbs; a smaller membrane, the propatagium, extending from the shoulder to the wrist (which had a long, thin bone called pteroid) and probably along the metacarpals, and another membrane between the legs (the uropatagium, which we will debate further). The propatagium and the uropatagium were simple membranes, but the main patagium, the cheiro/brachiopatagium, was very complex; it had several, extremely thin layers inside, composed of muscle fibers and collagen fibers (actinofibrils); supplying these layers were blood vessels, which unusually had loops on them. These fiber layers had several purposes; the muscle fibers imply the animal could extend and contract the membrane, thus changing the shape of the wing, making it thinner or wider. The actinofibrils were used to make the membrane more stiff, as mere skin would be too weak for flapped flight (bats do have wing fibers, though they are not as specialised as pterosaurian ones); being more concentrated on the distal part of the wing, they made the wing tips more rounded, as opposed to old reconstructions of pointy winged pterosaurs, as pointy wings would result in more falling than flying. As mentioned above, some pterodactyloids had air sacks on the wings; inflating them would probably help making the wing wider, though the main purpose was to pneumatise the forelimbs and perhaps supply the wing muscles with oxygen.
[Below, a picture of the Kitakyushu pterosaur by Matteo Bachin; note the rounded wing tips)

Shape of the wings

(On the right, Dimorphodon by Luis Rey; note the ankle attachement of the main patagia, the [cr]uropatagium dettached from the tail, and how it is thinner than the main patagium)
Having the wing tips rounded wasn't the only issue on pterosaur wing membranes; where in the legs were the main patagia attached? For years this has been a quite intense debate; generally the membrane is thought to be attached to the ankle, but some have suggested a knee attachment or not being attached to the hindlimbs at all. Fossil evidence seems to suggest an ankle attachment however, and since pterosaur wings could get thinner or wider it probably means there wasn't a lot of variation in attachment. Generally, pterosaur wings appear to have been quite thin according to fossils, but this could be post-mortem contraction of muscle fibers, and in any case pterosaurs could control their wing area. A big issue however is the shape of the uropatagium; some have suggested that it was attached to the tail as in bats, while others state it was independent from the tail and simply connected the legs (thus some reffer to it as "cruropatagium", heel membrane). This seems mostly of an issue on only primitive pterosaurs, as pterodactyloids clearly had a vestigial [cr]uropatagium, a simple membrane running along the legs; its possible this condition also existed in primitive pterosaurs, suggesting a mebrane independent from the tail and not connecting both legs.
As opposed to birds and bats which, having wings made of feathers and a membrane supported by more than one finger respectively, pterosaurian wings were quite limited in diversity of shapes. For instance, it'd be physically impossible to split the wing tips, as some birds do, and some flight techniques like static soaring would be difficult for pterosaurs to use. Still, wing shape diversity did occur; some forms had shorter and more rounded wings than others for instance.
Having the wing tips rounded wasn't the only issue on pterosaur wing membranes; where in the legs were the main patagia attached? For years this has been a quite intense debate; generally the membrane is thought to be attached to the ankle, but some have suggested a knee attachment or not being attached to the hindlimbs at all. Fossil evidence seems to suggest an ankle attachment however, and since pterosaur wings could get thinner or wider it probably means there wasn't a lot of variation in attachment. Generally, pterosaur wings appear to have been quite thin according to fossils, but this could be post-mortem contraction of muscle fibers, and in any case pterosaurs could control their wing area. A big issue however is the shape of the uropatagium; some have suggested that it was attached to the tail as in bats, while others state it was independent from the tail and simply connected the legs (thus some reffer to it as "cruropatagium", heel membrane). This seems mostly of an issue on only primitive pterosaurs, as pterodactyloids clearly had a vestigial [cr]uropatagium, a simple membrane running along the legs; its possible this condition also existed in primitive pterosaurs, suggesting a mebrane independent from the tail and not connecting both legs.
As opposed to birds and bats which, having wings made of feathers and a membrane supported by more than one finger respectively, pterosaurian wings were quite limited in diversity of shapes. For instance, it'd be physically impossible to split the wing tips, as some birds do, and some flight techniques like static soaring would be difficult for pterosaurs to use. Still, wing shape diversity did occur; some forms had shorter and more rounded wings than others for instance.
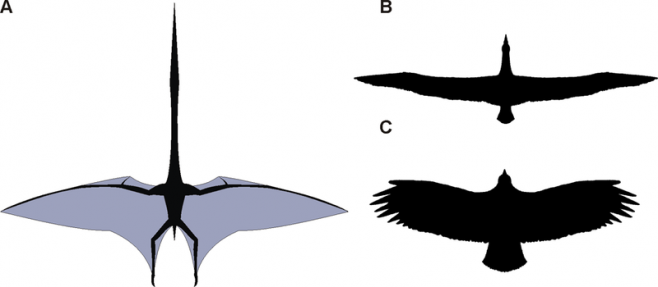
(A simple diagram comparing azhdarchid, condor and albatross wing shapes; living inland like condors, azhdarchids were physically incapable of efficient static soaring like them, so much like albatrosses they were probably more efficient gliders, being dynamic soarers)
Known flight types
With their wing limitations, pterosaurs seem less flexible than birds in flight styles, but in reality they were probably as diverse, using different techniques. From a bird biased view point, the main wing types expressed by pterosaurs would be high aspect ratio wings (as in seabirds), with some forms expressing wings akin to avian high speed wings (as in ducks and falcons) and elliptical wings (as in several forest birds like fowl and passerines), but considering the differences between pterosaurian and avian wings an exact comparation with these two wing types is difficult, though the animals that possessed them likely lived like their modern analogues. As far as it is known, dynamic soaring was the most common among large pterosaurs, even among forms that lived inland (dynamic soaring in modern birds seems to be most exclusive to seabirds); this is largely because static soaring, the style used by modern large inland flyers, was difficult for pterosaurs. It is possible that azhdarchids might have engaged in it, with their relatively shorter and possibly wider wings compared to other pterodactyloids, but this is uncertain. Other pterosaurs used different flight styles; dsungaripterids and tapejarids presumably flew like modern galliformes flew, using short flights with rapid wing beatings, while ctenochasmatoids probably flew like modern ducks and shorebirds, being relatively good and fast flyers.
Hovering in pterosaurs isn't very well studied; because pterosaurs couldn't rotate their forearms as in bats, they would have to do so bird style, by rotating their soulders, something difficult thanks to the fact that the wings were attached to the hindlimbs. In general, smaller pterosaurs probably achieved hovering more efficiently than larger ones, and based on the required power vs speed it was concluded that forms like Eudimorphodon, Pterodactylus, Rhamphorhynchus and Dorygnathus were more capable of hovering. However, since pterosaurs took a long time to grow it is possible that the juveniles of several large pterosaurs could hover. It seems that anurognathid pterosaurs might had been the better hovers of all pterosaurs, judging by several anatomical features like crests along their proximal humerus.
Hovering in pterosaurs isn't very well studied; because pterosaurs couldn't rotate their forearms as in bats, they would have to do so bird style, by rotating their soulders, something difficult thanks to the fact that the wings were attached to the hindlimbs. In general, smaller pterosaurs probably achieved hovering more efficiently than larger ones, and based on the required power vs speed it was concluded that forms like Eudimorphodon, Pterodactylus, Rhamphorhynchus and Dorygnathus were more capable of hovering. However, since pterosaurs took a long time to grow it is possible that the juveniles of several large pterosaurs could hover. It seems that anurognathid pterosaurs might had been the better hovers of all pterosaurs, judging by several anatomical features like crests along their proximal humerus.

(Image of Anurognathus by Mark Witton; these pterosaurs seem to have been the most efficient pterosaurs are hovering, and their flight style seems to have been similar to the one used by nightjars, owls and certain "large" bats, most a slow flight in search of prey)
Modern birds use their tail feathers for steering, while bats use their uropatagium (moving the legs); pterosaurs, logically, used their [cr]uropatagia as well. Pterodactyloids seem in general to have a less developed [cr]uropatagia than more primitive forms, which is generally consistent with their specialisation to more terrestrial niches, as a very developed [cr]uropatagium would get in the way when walking; its possible that ornithocheiroid pterosaurs had a more developed [cr]uropatagium, since they were generally more aerial.
Modern birds use their tail feathers for steering, while bats use their uropatagium (moving the legs); pterosaurs, logically, used their [cr]uropatagia as well. Pterodactyloids seem in general to have a less developed [cr]uropatagia than more primitive forms, which is generally consistent with their specialisation to more terrestrial niches, as a very developed [cr]uropatagium would get in the way when walking; its possible that ornithocheiroid pterosaurs had a more developed [cr]uropatagium, since they were generally more aerial.
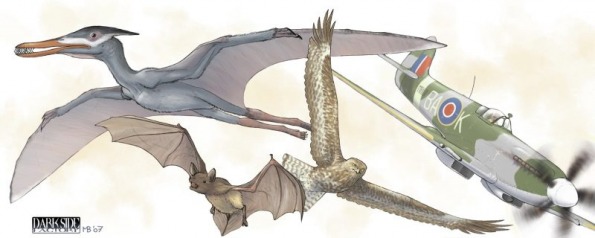
(Image of Cynorhamphus, bat, aeroplane and kite by Matteo Bachin (second "Dinosauri" book; published by "Jaca Book"); note the fact both the bat and the bird can split the wing end into several, something impossible for the pterosaur and the aeroplane [)
